Examples of close Euro-American connections:
Two Mediterranean species of Salix
A. K. Skvortsov
Euro-amerikanische verwandtschaftliche Beziehungen
zweier mediterraner Salix-Arten
Feddes Repertorium 1971, 82
(6): 407-420. Berlin
Translated from German by Irina Kadis
Translator's Note:
This article was written 36 years ago. We are providing the entire text, the way it
was published in German, except for the lists of herbarium specimens. According to the author himself,
this treatment is now partially outdated--particularly as regards the Californian willows.
However, the author has not changed his opinion regarding
S. tristis
and
S. humilis.
According to the contemporary treatment by G. Argus (1997), the American species mentioned in this
article are placed as follows:
S. lasiolepis and
S. irrorata to sect.
Mexicanae;
S. lemmonii and
S. geyeriana to sect.
Geyerianae.
As for
S. hookeriana, G. Argus has placed it together with
S. humilis var.
humilis and
S. humilis var.
tristis
in sect.
Cinerella, which corresponds to
sect.
Vetrix subsect.
Laeves in Skvortsov (1968).
This section as well includes the European species:
S. aurita,
S. caprea,
S cinerea.
The translator is grateful to Anett Hofmann
(University of Zurich)
for advice on some difficulties with German.
Abstract
The author shows a close relationship between
Salix salviifolia Brotero, a species distributed
across the Iberian Peninsula, and the eastern North American
S. tristis Ait. Both species, along with the
European
S. aurita L. and American
S. humilis Marsh., form a group of
closely related species within the subsect.
Laeves, sect.
Vetrix.
Salix pedicellata Desf., a
species with broad Mediterranean distribution, is related to the western
North American (mostly Californian)
S. lasiolepis Benth. The latter relationship, though it appears to
be more distanced than the former, is also apparent.
S. lasiolepis along with the closely related
S. hookeriana Barr. present a western
North American counterpart of the Old-World group of
S. pedicellata within the subsect.
Vulpinae. The available botanical evidence
supports the hypothesis of direct trans-Atlantic origin of these
connections rather than the explanation employing complicated migratory
processes across Beringia and East Asia.
Introduction
The phylogenetic relationships between willows of the Old and New
World so far rarely have been a subject of discussion. The only study was
undertaken in the 19th century by the renowned
Salix researcher N. J. Andersson (1858, 1868).
Andersson's conclusion was that "of 58 North American species, 24 are
identical with European ones; 24 belong to the same types, and only 10,
either western or Arctic forms, seem to be peculiar to this great
continent." (Andersson 1858). Though this researcher succeeded in pointing
out a number of valid connections, this conclusion of his today presents
only historic interest. The number of willow species known from North
America has grown more than twofold. Besides, the number of species common
with Europe is only half as large. If we take into account the five
European species that are widely cultivated in North America and
naturalized, then the total number of common species is still only 12.
From the perspective of contemporary knowledge, the majority of
Andersson's conclusions as regards species relationships appear incorrect.
(For example, he related
S. coulteri
and
S. lasiolepis to the European
S. daphnoides;
S. barklayi to
S. glauca;
S. phlebophylla to
S. retusa; he also considered
S. lucida to be a subspecies of
S. pentandra, etc.)
The relationships of the Old- and New-World willows have never been
revised since Andersson's time. Taxonomic research on the American and
European willows went on along two completely separate paths, totally
independently. C. K. Schneider (1920a, b) was studying Eurasian willows,
while his monographic description of the American species was not
completed. Most importantly, Schneider followed the American tradition in
his revision of the American willows and never succeeded to compare the
sections delimited within the New World with those described in the Old
World.
At present we have sufficient information only on the latest
(apparently Quaternary) Eurasiatic-American relationships within the genus
Salix. They are manifested first and
foremost among Arctic, Arctic-Alpine, and sub-Arctic species that are
common for both hemispheres (
Salix reticulata L.,
S. vestita
Pursh,
S. herbacea L.,
S. polaris Wahlenb.,
S. phlebophylla Anderss.,
S. rotundifolia Trautv.,
S. glauca L.,
S. ovalifolia
Trautv.,
S. arctica Pall.,
S. chamissonis Anderss.,
S. fuscescens Anderss.,
S. lanata L.,
S. alaxensis Coville,
S. pulchra Cham.,
S. sphenophylla A. Skv.
[1]
Specimens of the latter species from Alaska were identified by Hulten (1968) as
S. arctica
ssp.
torulosa
.
) and also in the boreal
S. bebbiana Sarg. Close affinity of the boreal American species
S. serissima Fern.,
S. planifolia Pursh, and
S. pedicellaris Pursh with the Eurasian
S. pentandra L.,
S. phylicifolia L., and
S. myrtilloides L. has been long known. The
author has highlighted (Skvortsov 1966) close connections between the
Siberian
S. boganidensis Trautv. and
S. erythrocarpa Kom. on one side and
the American
S. arbusculoides Anderss.
and
S. nivalis Hook. on the other
side. Hulten (1968) has noticed the presence of
S. hastata, an Old-World species, in Alaska.
These examples repeatedly refer to just Arctic and Arctic-Alpine species.
For the majority of American willows, the Old-World relations remain
unknown. Clarification of these connections presents the most important
and challenging goal of the
Salix
research.
This work is an attempt to approach the task through clarifying
relationships of two European willows,
Salix pedicellata Desf. and
S. salviifolia Brot., with American species. These links might
present interest not just for the willow research, but also to some extent
for the study of the Mediterranean flora and its connections.
Studied material and acknowledgements
This work is largely based on the study at the Herbarium of the
Botanic Institute in Leningrad (Curator Prof. I.T.Vassilczenko). Prof.
G.Moggi has kindly made it possible for me to handle extensive material
from the Herbarium of Florence University. Rich and valuable collections
of
S. salviifolia were received as
exchange from Dr. A.R.Pinto da Silva (Oeiras, Portugal). Some American
samples were provided by Prof. C.L.Hitchcook, Dr. Th.Crovello, and Dr.
P.H.Raven from the United States. A number of samples were studied by the
author in the Botanical Museum in Copenhagen (Director Dr. A.Skovsted).
Many thanks to all the colleagues.
Relations of Salix salviifolia
Brot.
The geographic area of this species is entirely within the
Mediterranean Floristic Area, provided that the Area is delimited in the
modern sense (see Meusel, Jäger, Weinert 1965). Yet
S. salviifolia is not purely Mediterranean or
sub-Mediterranean; even more appropriately it might be called
Mediterranean-Atlantic. This is true not only because it is distributed on
the Atlantic Coast, but also since it is more abundant in the western
areas of the Iberian Peninsula, in Portugal, than in the inner parts or
Mediterranean-Sea-facing areas of the peninsula.
S. salviifolia has not been found anywhere
outside the Iberian Peninsula.
The dense white-wooly tomentum of the lower leaf side is
characteristic for
S. salviifolia.
Indeed the species received its name because of this tomentum. This kind
of tomentum is found in species of sect.
Villosae (
S. lapponum L.,
S. alaxensis
Coville,
S. helvetica Vill.,
S. kryloviiE. Wolf,
S. candida Willd.) as
well as in sect.
Canae (
S. elaeagnos Scop.) Due to this similarity, some
authors (Andersson 1868, Pereira Coutinho 1899) were inclined to place
S. salviifolia close to the named
sections. Yet in the rest of its characters
S. salviifolia is rather different from the species of these
sections. It should without doubt be assigned to sect.
Vetrix (Vicioso 1951, Skvortsov 1968). More than
that, sections
Canae and
Villosae belong to a phylogenetic line that is
entirely different from that of sect.
Vetrix. The white villous trichomes of
S. salviifolia constitute one of many examples
of parallelism and convergence, which is so widespread in the genus
Salix (Skvortsov 1968). In other
species of sect.
Vetrix (such as, for
example,
S. pedicellata Desf. or
S. kuznetzowii Goerz), one can also
find specimens with nearly just as white and dense hairs.
Among Eurasian willow species, it is
S. aurita L. that is the closest to
S. salviifolia. From the first glance the two species look rather
unlike each other: one of them has narrow leaves covered with dense white
hairs, the other has broad leaves with sparse gray hairs. However, a
thorough morphological study of their organs reveals much
similarity.
One more species that has a lot in common with
S. salviifolia is the American
S. tristis Ait. Since Andersson's time (1859,
1868), most authors (including Schneider 1920a) mistakenly united this
willow with
S. sericea Marsh. and
S. petiolaris Sm. However, Ball (1947)
correctly placed
S. tristis in sect.
Vetrix (
Capreae). The position of this species can be
pinpointed even further. Together with
S. aurita and
S. salviifolia,
S. tristis forms a natural, wholesome
entity within subsect.
Laeves of sect.
Vetrix. As compared to the rest of
Vetrix representatives (such as
S. caprea L. or
S. cinerea L.), the members of this group
possess the following characters:
- shoots rather slender and short, more or less
covered with gray tomentum
- wood surface striated
- leaf petioles short; leaves mostly small, their
broadest part above the middle, revolute at margin, with rugose upper
surface, pronounced veins surfacing on underside, and more or less
tomentose pubescence
- flowering buds small, deltoid-ovoid
- catkins small, fruiting ones rather slender when
expanding, short in maturity; male catkins not larger than 25 x 16 mm
in S. aurita; in S. salviifolia and S. tristis even smaller than that (whereas
catkins of, say, S. caprea often
become as large as 50 x 27 mm)
- bracts small, mostly obtuse and only seldom entirely
black; they are mostly colored purple-brown or entirely pale, with
scarcely blackish tips
- anthers small when dry: 0.5-0.6 mm long, as compared to 0.7-1.1 mm in
S. caprea-S. cinerea group)
- styles and stigmas rather short (stigmas about
0.2-0.4 mm long)
- S. aurita and
S. tristis are known to be
restricted mostly to poor, acid soil; we don't know for sure if this
is true for S. salviifolia;
however, it is quite probable.
It is not easy to decide to which of the two related species
S. salviifolia is closer. Its
pronounced wood striation, the shape of buds and stipules, as well as male
catkin morphology make it closely resemble
S. aurita. In its leaf shape and pubescence as well as in
morphology of developing fruiting catkins,
S. salviifolia has much in common with
S. tristis. As regards other characters, such as petiole length,
leaf blade size, degree of stipule development, or catkin size,
S. salviifolia occupies a position intermediate between
S. aurita and
S. tristis. In comparison with the other two species' characters,
those of
S. salviifolia can be
presented as follows:
|
S. salviifolia
|
S. aurita and
S. tristis
|
|
A tall shrub or a small tree. Pubescence on leaf
undersides very much pronounced, mostly white.
|
Mostly tall or small shrubs. Pubescence on leaf
undersides mostly gray, often disappearing in mature leaves.
|
|
S. salviifolia
|
S. aurita
|
|
Leaves 4 to 7 times as long as broad.
|
Leaves 1.5 to 3 times as long as broad.
|
|
Leaf margin mostly denticulate or entire.
|
Leaf margin mostly irregular, undulate, coarsely dentate.
|
|
Leaf
apex acute or more or less obtuse, straight.
|
Leaf apex rounded
or abruptly attenuating in a short, often crooked
point.
|
|
S. salviifolia
|
S. tristis
|
|
Wood striae mostly pronounced, long, multiple.
|
Wood striae small, sparse.
|
|
Stipules broad, inequilateral, obtuse.
|
Stipules
smaller, often nearly lanceolate and nearly equilateral,
acute.
|
|
Floriferous buds broad deltoid-ovoid.
|
Floriferous buds narrower: up to
oblong-ovoid.
|
|
Male
catkins oblong-cylindric, 15-25 x 10-14 mm.
|
Male catkins oblong to nearly globose,
6-12 x 7-10 mm.
|
S. tristis is distributed in the
eastern US from Massachusetts to northern Florida, westward to North
Dakota and Nebraska.
In American literature
S. humilis Marsh. is commonly positioned close to
S. tristis. According to such researchers as
Griggs (1905), Schneider (1920a), Argus (1964),
S. humilis and
S. tristis are just two extremes connected through a number of
intermediate forms. This concept might be not right. Morphologic
boundaries between species and diagnostic characters definitely require
further elaboration, yet the treatment of
S. tristis as a distinct species already appears to be sufficient.
S. tristis differs from
S. humilis morphologically, ecologically, and
also geographically.
S. humilis may be
considered as yet another, fourth member of our species group. The
majority of diagnostic characters for this group that have been described
here above also fit
S. humilis. At the
same time,
S. humilis, due to some of
its characters, constitutes the link connecting the group of
S. aurita-
S. salviifolia-
S. tristis with
S. caprea.
Leaves of
S. humilis, glabrous
on the upper side, covered with spreading trichomes on the lower side,
with their relatively long (4-12 mm) petioles, resemble
S. caprea leaves.
Some samples of
S. humilis with
leafy branches generally look hardly different from small-leaf forms of
S. caprea. This is particularly true
for northern plants, the so-called var.
keweenawensis
Farwell, such as, for example, Bartram & Long 23745 from Nova Scotia,
Garton 6305 from Ontario, or Jesup 37 from Massachusetts. In
S. humilis anthers are larger than in
S. tristis. Also,
S. humilis has a larger range that is shifted more to the north in
comparison with the range of
S. tristis.
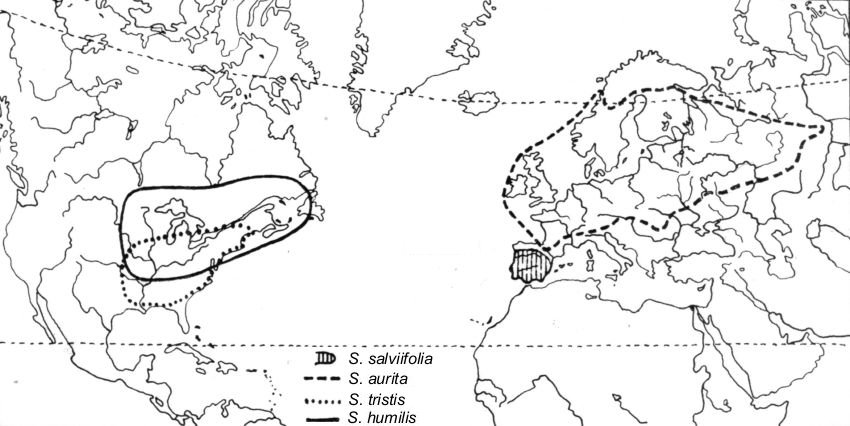
As it is obvious from the enclosed map,
the entire group of species that we are talking about can
be considered amphi-Atlantic. It is not represented anywhere in Asia
(except for a tiny marginal part of
S. aurita area in the Ural Mts., within the 500-mm isopluvial
line.) This group is as well absent from western North America. Hulten
(1958) has explained similar amphi-Atlantic distribution patterns by
assuming connections in the past via Beringia and East Asia. However, in
East Asia, which is generally so rich in species including primitive and
relict groups of the genus
Salix, not
a single relative of the species in question has been found. If we
accepted a possibility of direct trans-Atlantic Euro-American connections
during the period from the beginning to mid-Neogen, then we could better
explain the distribution patterns observed at present.
[List of observed herbarium specimens--S. salviifolia and S. tristis--omitted]
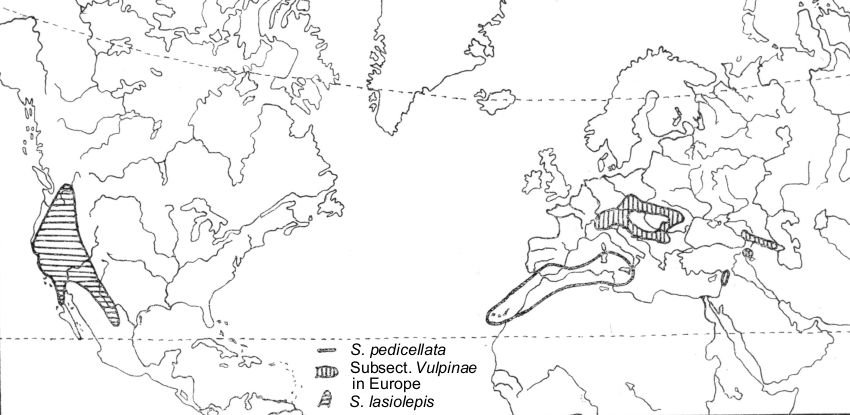
Trans-Atlantic connections of S. pedicellata Desf.
The author believes that
S. pedicellata belongs to sect.
Vetrix, subsect.
Vulpinae together with such species as
S. vulpina Anderss.,
S. silesiaca Willd.,
S. appendiculata Vill.,
S. caucasica Anderss. The close relation between the latter three
species and
S. pedicellata is
apparent, while its connection with the Far Eastern
S. vulpina is much less obvious. Together with
some other Japanese-Korean willows,
S. vulpina forms a tightly knit group within subsect.
Vulpinae.
Differently from
S. salviifolia,
S. pedicellata is a typical
Mediterranean species, according to its range. Its complete area lies
within western Mediterranean area: Sicily, Sardinia, western Spain, and
North Africa. It penetrates the eastern Mediterranean only in Lebanon and
the adjacent parts of Turkey (and probably also on some of the Aegean Is.
Subsp.
canariensis (Chr. Smith) A. Skv. is
distributed on Canary Is. and Madeira.
When looking for
S. pedicellata
relations among North American willows, we stop at
S. lasiolepis Benth. This species is widespread
in western North America from southern British Columbia to western Texas
and northern Mexico.
S. lasiolepis is
particularly frequent in California, where it is one of the most common
willows. Characters shared by
S. lasiolepis and
S. pedicellata can be summarized as follows:
-
Both species are distributed from the sea level to intermediate
and even high elevations in the mountains,
S pedicellata ascending to 800 m on Madeira and Sardinia, to
1,100 m in Lebanon, to 2,200-2,400 m in North Africa;
S. lasiolepis reaching 1,500-1,600 m in
California, 2,100-2,200 m in Mexico.
-
Both have a vast ecologic range: from alluvial habitats to
wetlands.
- Both willows are tall shrubs or trees (10-12 m tall)
with broad crowns and often rather thick trunks.
- Shoots mostly colored somewhat dirty tawny, more or
less covered with short gray felted pubescence.
-
Floriferous buds deltoid-ellipsoid, 4-7 mm long, with pronounced
lateral carinas, their tips straight (or bent toward the branch,
rarely somewhat recurved), flattened on the adaxial side.
- Stipules mostly well developed, broad, oblique,
semi-cordate, rather asymmetric.
- Petiole thick, convex above (round on
cross-section), mostly downy.
- Leaf blade shape similar to that in S. cinerea or S. atrocinerea, with the broadest part mostly above the
middle. Leaf apex obtuse or short-pointed.
- Margins in mature leaves revolute,
with more or less sparse, irregular teeth.
Glands submarginal.
- Lower leaf surface more or less covered with
pubescence, first spreading, then becoming felted; lower surface often
glabrous. Venation prominent on underside of mature leaves.
- Catkins precocious (emerging earlier than leaves);
in warm areas expanding in January-February, upright, medium-sized,
relatively slender, oblong-cylindric. Bracts rather small, hairy, with
blackish tips only or completely without black parts.
- Stamen filaments glabrous, rather short; anthers on
average 0.5-0.7 mm long.
- Ovaries medium-sized, long-stipitate, conspicuously
protruding from the bract pubescence already at the time of flowering,
mostly glabrous, gradually attenuating into short styles. Styles pass
into rather small two-parted stigmas.
It's important to note that
S. pedicellata, as regards some of its characters (such as
distribution across altitudes and latitudes, habit, floriferous bud
morphology), is closer to
S. lasiolepis than any of the European-Caucasian related species.
As for differences between
S. pedicellata and
S. lasiolepis, they may be summarized as follows:
|
S. pedicellata
|
S. lasiolepis
|
|
pronounced wood striation under bark
|
wood striation absent
|
|
leaf margin often with large teeth (like in S. cinerea)
|
leaf-margin denticles small
|
|
leaf blades rather smooth and thin, upper side opaque
|
leaf blades more or less firm, coriaceous, their
upper side smooth, more or less lustrous
|
|
bracts usually longer than broad, not truncate, with
straight hairs, which overtop bract tips by 1.0-1.5 (-2.0)
mm
|
bracts as broad as long or even broader, rounded or
truncate at tips, with rather dense, somewhat wavy hairs, which
overtop bract tips by 0.3-0.8 (-1.3) mm
|
|
stamen filaments free
|
stamen filaments often connate from
base
|
|
capsule stipes 1.5-4.5 mm long
|
capsule stipes 0.8-2.0 mm long
|
|
style 0.2-0.4 mm long
|
style length varies from 0.2 to 0.8 mm
|
S. pedicellata ssp.
canariensis is different from its continental
counterpart in weaker wood striation, longer leaves with more lustrous
upper surface, and shorter capsule stipes. Considering this character
expression, we can place this subspecies even closer to
S. lasiolepis. Besides, even within
S. pedicellata sensu stricto one can find
specimens in which wood striae are nearly absent, such as, for example,
Desfontaines' specimen preserved in LE. On the other hand, in any large
sample of
S. lasiolepis, there may be
found specimens with longer and straighter bract hairs.
In the American literature,
S. lasiolepis is usually placed in sect.
Cordatae Barr. The reason for that is,
apparently, the similarity between
S. lasiolepis and members of
Cordatae in their large-sized, glabrous,
stipitate, more or less pointed capsules. However, this type of capsules
is not unique for
Cordatae. Very
similar capsules are also found in
S. pedicellata,
S. silesiaca,
S. caucasica, the east-Asian
S. vulpina, as well as in
S. japonica Thunb.
and
S. shiraii Seemen and some members of sect.
Helix. As for the rest of the characters,
members of sect.
Cordatae are
different from
S. pedicellata in
nearly every other respect. The true members of sect.
Cordatae, both American and Eurasian,
[2]
The Eurasian sect.
Hastatae
Kern. certainly has to be identified with
Cordatae; the name
Cordatae
has the priority.
all have either straight boreal or even sub-Arctic
distribution. In southern parts of their areas, they are mostly restricted
to montane forests or
sub-Alpine and Alpine belt. The species of sect.
Cordatae are drastically different from
S. lasiolepis in the following
characters:
- young branches mostly bright colored
- floriferous buds laterally rounded, without
carinas
- stipules equilateral or sub-equilateral
- petioles flat or convex on the upper side
- leaf blades with veins slightly conspicuous or
inconspicuous on the underside
- leaf margins barely
revolute, regularly and densely denticulate
- glands truly marginal
Due to these significant differences,
S. lasiolepis should be removed from sect.
Cordatae and placed in sect.
Vetrix, subsect.
Vulpinae.
According to Schneider (1920b) and Ball (1961), a species closely
related to
S. lasiolepis is
S. irrorata Anderss., which was also placed by
these authors in sect.
Cordatae. This
opinion is completely unacceptable.
In accordance with the structure of its vegetative organs as well as
catkins,
S. irrorata has even less in
common with the species of
Cordatae
than
S. lasiolepis. However,
S. irrorata does not have much in common with
sect.
Vetrix, either. Perhaps we
should consider placing
S. irrorata
together with
S. geyeriana Anderss.
and
S. lemmonii Bebb.
S. tracyi described by Ball
(1934) in my opinion is merely a poor (probably overtopped) form of
S. lasiolepis with slenderer twigs and
thinner, scarcely pubescent leaves. It flowers later and much less
abundantly than
S. lasiolepis. When
the entire variability range of
S. lasiolepis is taken into account, differences between
S. lasiolepis and
S. tracyi, as described by Ball (1934), appear not to be
convincing enough.
Among the American willows, it is
S. hookeriana Barr. that is the closest to
S. lasiolepis. This was first noticed by
Crovello (1968). Before that
S. hookeriana had been placed together with
S. lanata,
S. barratiana, and
S. alaxensis. In comparison with
S. lasiolepis,
S. hookeriana
has thicker branches, larger buds, leaves, catkins, anthers, capsules, and
stigmas; capsules are only very short-stipitate. All these differences,
however, are purely quantitative; by no means they are contradicting the
close affinity of the two species. Apparently, the
S. lasiolepis-
S. hookeriana group should be considered a special
western-American entity within subsect.
Vulpinae.
Due to lack of herbarium material, I could not reliably clarify the
taxonomic value and position of
S. piperi Bebb. It is quite possible that this is merely a form of
S. hookeriana with weakly developed
pubescence, which was described as a species similarly to
S. tracyi, a form of
S. lasiolepis.
East Asian members of subsect.
Vulpinae --
S. vulpina and the closely related
S. sieboldiana Blume and
S. buergeriana Miq., in accordance with their morphological
characteristics, cannot be considered a link between the groups of
S. lasiolepis-
S. hookeriana and
S. pedicellata-
S. caucasica.
Similarly to the case of
S. salviifolia, the hypothesis employing a possibility of direct
trans-Atlantic early-Neogenic connections appears to be more convincing
than the more complicated assumption of a connection via Beringia and
Asia.
[List of observed herbarium specimens--S. pedicellata, S. lasiolepis,
and S. hookeriana--omitted]
References
Andersson, N. J.1858. Salices boreali-americanae.
--
Proceed. Amer. Acad. 4, Separatum: 1-32
Andersson, N. J.1868. Salix. In: De Candolle, A.P.
Prodromus systematis naturalis 16 (2): 190-323
Argus, G. W.1964. Preliminary reports on the flora of Wisconsin 51. The genus
Salix.
--
Proceed. Wisconsin Acad. Sci. 53: 217-272
Ball, C. R.1934. New or little known West American willows.
--
Univ. Calif. Publ. Bot. 17, N 14: 399-434
Ball, C.R.1947. Studying willows or making new sections in the genus Salix.
--
Rhodora 49: 37-49
Ball, C.R.1961. Salix. In: Lundell, C.L., Flora of Texas 3, part 6:
369-392
Crovello, Th.G.1968. A numerical taxonomic study of the genus Salix, section Sitchensis
--
Univ. Calif. Publ. Bot. 44: 1-61
Griggs, R. F.1905. The willows of Ohio
--
Proceed. Ohio Acad. Sci. 4: 256-314
Hulten, E.1958. The Amphiatlantic plants. Stockholm
Hulten, E.1968. Flora of Alaska and neighboring territories. Stanford
Meusel, H., Jäger, E., and Weinert, E.1965. Vergleichende Chorologie der zentraleuropaischen Flora. Jena
Pereira Coutinho, A. X.1899. Subsidio para o estudo das Salicaceaes do Portugal.
--
Bol. Soc. Broter. 16: 5-34
Schneider, C. K.1920a. Notes on American willows. IX
--
J. Arnold Arbor. 2: 1-25
Schneider, C. K.1920b. Notes on American willows. XI
--
J. Arnold Arbor. 2: 185-204
Skvortsov, A. K.1968. Ivy SSSR. Sistematicheskiy i geograficheskiy obzor [Willows of
the USSR. Taxonomic and geographic revision.] Moscow: Nauka. 262 pp.
Skvortsov, A. K.1966. Salix. In: Arkticheskaya flora SSSR [Arctic Flora of the USSR] 5.
Leningrad: Nauka. Pp. 7-118
Vicioso, C.1951. Salicaceas de Espana. Madrid
NOTES
Translation I.Kadis
31 March 2007