Present distribution and probable primary range
of brittle
willow (Salix fragilis L.)
A. K. Skvortsov
Problemy biogeotsenologii, geobotaniki i botanicheskoy
geografii. Leningrad, Nauka, 1973, pp. 263-280
Translation: Irina Kadis
Introduction
It was Andersson (1868: 209) who first came up with the idea that
Salix fragilis L. was not truly wild
and autochthonous in Europe and that it probably originated in southwest
Asia. Andersson considered another willow that he had described from
southwest Asia, S. australior
Anderss., to be a wild ancestor race of S. fragilis. This provided grounds for his hypothesis about the
Asiatic origin of S. fragilis.
However, later on it became clear that Andersson was mistaken when he
considered S. australior to be closely
related to S. fragilis. S. australior (priority name S. excelsa Gmel.) rather belongs to the cycle of
S. alba L. Therefore, the hypothesis
about brittle willow's Asiatic origin was abandoned. No further studies
were conducted in order to find out whether it was aboriginal in
Europe.
While completing the taxonomical revision of the willows distributed
within the USSR and surrounding countries, I, too, had to face the problem
with S. fragilis. The analysis of all
available material has brought me to the conclusion that S. fragilis is adventive in the USSR and Europe.
Its primary range is within northern and northeastern Asia Minor. Though
my reasoning behind the judgement on the motherland of S. fragilis is
entirely different from Andersson's, I arrived to a very similar
conclusion. Even though I have already presented it in the review of the
willows of the Caucasus and Asia Minor and also in the review of the USSR
willows (Skvortsov 1966: 116-117; Skvortsov 1968: 68, 111-113.), detailed
arguments were beyond the scope of these works. The purpose of the article
is to provide this argumentation.
I used the herbarium material at most major depositories in this
country: herbaria of the botanical institutes in Leningrad, Kiev, Tbilisi,
Baku, and Yerevan; Moscow and Tomsk university herbaria; and also the
Herbarium of the Natural History Museum in Lvov. Of foreign depositories,
I examined all the material at the Prague National Museum and collections
from Asia Minor at the Royal Botanic Garden in Edinburgh. I express my
cordial gratitude to all the colleagues who run collections at these
institutions. I wish to thank Dr. E.L. Swann (Norfolk, England) and Dr.
A.R. Pinto da Silva (Oeiras, Portugal) for some valuable material they
mailed for the study. I am indebted to Dr. Neumann (Vienna) for his
important comments on the distribution of S. fragilis in Western Europe.
The late E.I. Steinberg provided translations of Estonian texts.
I have conducted field observations primarily in Moscow, Smolensk,
Kaluga, and Tula oblast's. Later on I included the vicinity of Leningrad,
the Carpathians, and Volgograd Oblast. Additionally, I conducted some
observations at a lesser scale in Pskov, Novgorod, Ryazan, Voronezh,
Saratov, Belgorod, and Kiev oblast's, and also in Latvia and central
German Democratic Republic. Literature data were extensively used as
well.
I. Brittle willow in central European Russia
Here is what Tsinger (1886: 393) said about S. fragilis in his
Anthology of Knowledge:
"It is very common everywhere along rivers, around ponds, at dams,
near dwellings, and at roadsides; for Tver Government it is listed only as
cultivated. It probably becomes rare in the southeast, in the proximity of
the Volga banks." This description was inherited by the early editions of
Mayevskiy's Flora of Central Russia (1892: 413; 1895: 435; 1902: 469). The
note regarding Tver Government found in Tsinger (1886) as well as the
first three editions of Mayevskiy's Flora, had been originally made by
Bakunin (1879: 348).
Petunnikov (1901: 37-39), citing mostly F.A. Teploukhov's opinion
(Teploukhov apparently never published the results of his willow studies),
stated that the majority of samples treated as S. fragilis within Moscow Government were actually the so-called
S. viridis Fr., i.e., hybrids of
S. fragilis and S. alba. At the same time, according to
Petunnikov, the true, non-hybrid S. fragilis
was rather rare in Moscow Government. Remarkably, all of
non-hybrid specimens of S. fragilis
that Petunnikov was aware of turned out to be staminate. Siuzev (1901) was
another author citing Teploukhov's opinion on the hybrid nature of most
S. fragilis in this country. He
provided quotations from a letter by Teploukhov, which even indicate that
Teploukhov probably had doubts about the aboriginal nature of S. fragilis. At the same time, Siuzev argued
that indeed he himself had found pistillate specimens of S. fragilis on
the Oka River in Moscow Government, and therefore Teploukhov finally
agreed that S. fragilis was a native
species in the flora of this country.
Syreshchikov (1907: 15, 19; 1927: 98, 99) noted in support of
Petunnikov's view that S. viridis was
common in Moscow Government, while S. fragilis was rare. The Flora of Central Russia by Mayevskiy,
starting from edition 4 (1912: 500), also says that "crosses with
S. alba occur more often than the
typical" S. fragilis; while the note
about Tver Government vanishes from the text. Starting from that time, the
suggestion that S. fragilis might be
not native in central European Russia is not repeated in the
literature anymore. It is not found in the regional "floras" of Vladimir
and Kaluga (Flerov 1902, 1912), Tula (Rozen 1916), or Kalinin (Nevskiy
1952).
My own observations in central European Russia completely support
the conclusions of the earlier authors about domination of hybrids
S. fragilis x S. alba over "pure" S. fragilis. Though the two species belong to the same section,
they have a number of quite distinct differences. Therefore, recognition
of hybrids should not be just an intuitive procedure based solely on
inspiration, asâalas!âit often happens. Instead it can be
based on distinct morphological criteria.
The diagnostic characters of the two species are superposed here
below.
|
S. fragilis
|
S. alba
|
|
Trees developing in the open produce
low, nearly spherical crowns; large oblique limbs originate at a
fairly low level
|
Crowns of trees developing in the open
mostly elongate; trunks upright, tall; limbs relatively
short
|
|
Within old part of crown, 1-3-year-old
branches never pendulous, spreading at 60-90° angles, breaking off easily when gently
pressed
|
Within old part of crown, 1-3-year-old
branches often pendulous, growing at angles less than 60°, not as
brittle
|
|
It is only current-year branchlets,
particularly suckers, that often become pigmented red or rich
brown; older branches within old crown uniformly yellowish-gray or
nearly ivory, occasionally with some orange tint or reddish
spots
|
Current-year branchlets mostly green;
older branches becoming rather bright reddish or brownish,
pigmentation more pronounced on the lighted side
|
|
Current-year branchlets may be
puberulent; older branches always completely
glabrous
|
Current-year branchlets always
pubescent; older branches mostly pubescent, at least in distal
parts
|
|
Buds during late fall and winter colored
same as branches; apices blackening due to bud scale dieback. Buds
ovoid to lanceoloid, more or less acutish, abaxially convex or
more or less carinate; slightly convex or sometimes also carinate
on adaxial side
|
Buds during the fall and winter colored
same as branches or brighter, not blackening,
oblong lanceoloid, mostly blunt, abaxially
somewhat convex; mostly completely flat adaxially
|
|
Outermost leaf primordium in bud mostly
pale or grayish brown, broad, nearly round when spread, with
several parallel veins originating from base. Catkin primordia in
floriferous buds densely sericeous
|
Outermost leaf primordium mostly
reddish, oval, its venation pinnate. Catkin primordia in
floriferous buds puberulent, greenish; imbricate bracts form
distinct pattern visible through pubescence
|
|
Young leaves in spring light
yellowish-green; mature foliage dark green, without bluish
tint
|
Young leaves more or less deep
bluish-green; mature leaves pale bluish-green or
sericeous
|
|
Stipules obliquely semi-cordate,
distinctly inequilateral
|
Stipules narrowly lanceolate,
linear-subulate, subequilateral, rarely somewhat
falcate
|
|
Petioles 7-15 mm long; petiolar glands
pronounced in all leaves on the branch, except for proximal ones
|
Petioles 2-8 mm long; petiolar glands
pronounced mostly only in distal leaves and on suckering
shoots
|
|
Young leaves mostly completely glabrous,
occasionally sparsely hairy; mature leaves glabrous
|
Young leaves delicately sericeous;
mature leaves also more or less silvery pubescent
|
|
Leaf blade broadly cuneate or rounded at
base; reticulation network very conspicuous on underside. Margin
dentation rather coarse, denticles mostly bent or rounded. Stomata
on upper leaf surface mostly sparse or wanting
|
Leaf blade mostly cuneate at base; only
major lateral veins are visible on underside, reticulation
wanting; margin minutely serrulate, denticles mostly acute.
Stomata dense on upper leaf surface
|
|
Catkins spread nearly at right angle
from branch. All flowers open simultaneously in staminate
catkins
|
Catkins positioned at acute angle on
branch. Proximal flowers in staminate catkins open earlier than
distal
|
|
Bracts 0.8-1.5 x 0.5-0.8 mm, covered
with straight trichomes that extend by 0.8-2.0 mm beyond bract
margin. Bracts remain pale on drying
|
Bracts 1.2-2.8 x 0.7-1.0 mm,
puberulent; trichomes extend by 0.2-0.6 mm beyond bract margin.
Bracts turn rusty-brown on drying
|
|
Filaments pubescent only at base (along
â
to ž of their length); dry anthers 0.5 (0.6) mm
long
|
Filaments pubescent up to ž-Â― of their
length; dry anthers (0.5) 0.6-0.7 mm long
|
Hybrids occupy and intermediate position, being closer to one or the
other parent as regards characters expression. This variability is
understandable, as we deal not only with F
1
generation, but various products of segregation formed as a result of
reciprocal crosses with parental species as well as crosses within
F
1. Heterosis is typical for the majority of
hybrids. Usually hybrids grow faster and produce more robust branches than
parents
[1]Some authors, for example, Nazarov (1949: 54) have had an
impression that pistillate plants of S. fragilis tend to grow faster than staminate. Such
observations without doubt have to be attributed to comparisons of
hybrid pistillate and non-hybrid staminate specimens..
There also exist hybrids of S. fragilis with S. pentandra
L. and probably with S. triandra L.;
however, those are rare in central European Russia and present no interest
for this discussion.
Differently from Siuzev's experience, my search did not yield any
clearly non-hybrid pistillate S. fragilis around the Oka River or elsewhere in central European
Russia. Trees displaying typical species characteristics were invariably
staminate. More than that, the herbarium search within material from this
area yielded the same result: not a single pistillate specimen was found
without hybrid traits. One could speculate that we are dealing here with a
case of sexual dimorphism, i.e., pistillate plants are characteristically
different from staminate ones in many respects. However, this assumption
does not stand under scrutiny, as pistillate S. fragilis are found in other parts of the area, where they are
similar to staminate plants in every respect (except for their sex).
Siuzev's statement about presence of "pure" pistillate S. fragilis at the Oka River was apparently
based on insufficient character analysis.
The predominance of hybrids along with complete absence of
non-hybrid pistillate specimens in such a wide-ranging species as
S. fragilis within the central
European Russia presents a puzzle that needs explanation. It is Bakunin's
opinion that comes to mind first. He believed that within Tver Government
all of S. fragilis was just
cultivated. However, brittle willow in Moscow, Kaluga, Smolensk, Tula, and
Ryazan oblast's is commonly found in habitats were it sure could not have
been planted. In the aftermath of a storm or during a windy day,
multitudes of broken twigs are found around every old brittle willow. Many
branchlets are carried away by wind or running water and root in 2-3
weeks, if they happen to be in favorable moisture situations. Thus
S. fragilis is a spontaneous species
undoubtedly belonging to the wild flora. However, does it also happen to
be indigenous? In order to answer this question, it is necessary to
consider habitats taken by the species and decide whether there exists a
niche for it anywhere in undisturbed nature.
For solving a problem of this type, historic records also might be
of some value; however, in this case chances of finding positive evidence
are more than slim. Besides, historic evidence can be only relatively
meaningful. If we cannot provide documentation describing the introduction
of potatoes to Kamchatka, that does not necessarily mean that potatoes are
native there. On the other hand, even though we know for sure that grapes
have been cultivated in Tajikistan for ages and even millennia, we still
cannot exclude a possibility for wild species to exist there. Whether or
not we discover indications of oriental plane cultivation in the Varzob or
Tsav river valleys, the outcome is not going to alter the fact that plane
is a member of the native flora in Armenia. In all such cases, we should
base our conclusions first and foremost on biological observations.
Outside truly secondary, man-made habitats, such as roadsides,
ditches, or residential lots, S. fragilis has been found along streams and rivers, in damp
depressions, and in small spring-fed fens. The most common willows on
river banks in this country are S. viminalis L., S. dasyclados
Wimm., S. triandra L., and S. alba L. There is no doubt that these four are
native to the area. Whenever there appears a freshly deposited alluvium
patch in a river valley, it is promptly taken by these species. It is
mostly S. viminalis that occupies
sandy deposits right along the water margin; S. triandra and S. dasyclados
both require silt deposits and thus settle particularly along slow creeks
or surface run-off depressions; still more often all the three species
form mixed thickets in various proportions.
As for S. alba, it favors
sandy-silty deposits that are formed during large floods at high levels,
which remain dry during most of the year. Depressions that remain moist
for a long time after the flood are particularly favorable. Thus it forms
pure or nearly pure stands in large river valleys, usually producing small
and dense, even-aged groves extended along river beds. Most of these
groves have been clear-cut; however, I have found some along the Oka and
Protva that were quite intact. When S. alba happens to grow along small streams, it is usually only
solitary and, as a rule, occurs not far from some large river
valley.
S. alba, S. viminalis, S. dasyclados, and S. triandra
are consistently found at suitable habitats in semicultural landscapes as
well as in those only moderately affected by human activities. The less
the landscape has been altered by humans, the more the original ecological
preferences of each species are pronounced, and the more often we
encounter good seed reproduction at appropriate habitats.
In contrast to the species named above, S. fragilis never forms continuous single-aged groves in large
river valleys, particularly along the Oka. We always find only scattered
solitary trees of different ages that randomly occur in different parts of
river valleys, most often within the floodplain, around oxbow lakes, along
small tributaries, or in erosional depressions formed by floods. These
habitats are also preferred by S. alba, which naturally grows there. Healthy natural groves of
S. alba never contain any S. fragilis, even as a minor admixture. In
undisturbed situations, all habitats suitable for S. alba become colonized by S. alba only, even though there are fruiting
hybrids S. alba x S. fragilis growing in close proximity. These
hybrids along with pure S. fragilis
are encountered only in habitats fairly disturbed by humans. Hence, when
natural habitats favorable for S. alba
are available, hybridization between S. alba and S. fragilis,
however extensive it is, has no effect on the integrity of S. alba, its ecological and morphological
stability as a species. It is only S. fragilis that is subject to disruption and erosion as a
species.
During my tours of the major rivers (Oka, Moskva, Protva, Ugra,
Osetr, and Upper Dnieper), I have never encountered any large-scale
reproduction of S. fragilis or its
hybrids. Consequently, we should admit that within the major river valleys
in this country there are no habitats where S. fragilis could exist prior to human disturbance.
Yet what about those small rivers, brooks, and spring-fed fens?
Perhaps S. fragilis could be a natural
part of plant communities or successions there, could not it? All of our
native forest trees, including those that never dominate climax formations
or major successional stages, are constantly found (even though they might
be scarce and depressed) in the oldest, minimally disturbed forests. This
is not the case with S. fragilis. It
is not consistently present in any climax communities, even as a rare
species, neither found regularly at any particular successional stage.
Stream banks and moist bottoms of depressions in this country used to be
naturally covered by black alder groves and stream-bank spruce
associations. We cannot imagine a light-loving tree with a broad low
crown, such as S. fragilis, growing
amidst a dense spruce forest or black-alder grove. It could not even
temporarily be present during the natural regeneration stages of these
communities.
Openings in black-alder groves and spruce forests that occur due to
wind or insect damage never affect the forest environment to such a
significant extent that it ceases being a forest; and S. fragilis never trespasses forest habitats, as
far as we know from observations. Whenever moist forest grounds start
receiving more light, tall herbs would conquer the spot along with
S. caprea L., S. aurita L., and sometimes S. cinerea L., yet never S. fragilis. It is only present in the
situations when light has been available for a long time and forest
character of the habitat has been long lost to a large extent. It is only
then that S. fragilis may settle,
self-dispersed not from seed, but rather from broken twigs brought in by
wind or water.
We should conclude that within the natural vegetation cover of
central European Russia there is no niche for S. fragilis to occupy. We thus have to acknowledge that brittle
willow is an adventive plant in this area.
II. Brittle willow in other regions of the USSR (excluding the
Caucasus)
Many "floras" and reviews (Wolf 1900: 21; Seemen 1908-1910: 70;
Nazarov 1936: 203; Pravdin 1951: 168; Rechinger 1957: 67, and others)
inform that S. fragilis is distributed
across the entire European territory of the USSR, except for the extreme
north, and that its range also extends to West Siberia and the Altai Mts.;
some believe that this willow even reaches the Lake Baikal (Penkovskiy
1901: 62). However, herbarium data signify that S. fragilis (together with its hybrids) hardly
crosses the Volga. East of the Volga, it is represented only by
some solitary findings. I have seen five specimens from Siberia and
Kazakhstan (Biysk, Alma-Ata, Akmolinsk, Karaganda, and Telmanovo in the
vicinity of Tselinograd) and some very scarce from the southern Urals and
Bashkiriya. A single concrete reference to S. fragilis from the Altai Mts. (Edzhigan ValleyâKrylov) has
turned out to be a misidentification (Skvortsov 1956).
The notion of predominance of hybrids S. fragilis x S. alba over
"pure" S. fragilis was extrapolated
over the entire range of this species within the USSR by Nazarov (1936) in
the Flora of the USSR. However, a detailed herbarium study and
differential mapping suggest a more intricate and interesting relationship
between hybrid and non-hybrid S. fragilis. Remarkably, all of the named five specimens from
Siberia and Kazakhstan happened to be hybrids. Specimens from Alma-Ata and
Biysk are known to be collected from cultivated plants. The remaining
three samples from Kazakhstan originate from regions where S. alba is not distributed, which makes the
formation of autochthonous hybrids impossible. Apparently, these were also
collected from cultivated plants.
Non-hybrid S. fragilis was never
ever collected from the Urals or Upper Vyatka Basin, neither from Volga
Uplands. Hybrids of S. fragilis are
not infrequent in Saratov and Volgograd oblast's, even away from human
dwellings, where they appear to be growing naturally. However, a more
careful examination always shows they have been planted. Apparently, in
the Volga Basin there is no current advancement of S. fragilis hybrids, neither by seed
dissemination, nor even from vegetative parts.
Further herbarium study has shown that the entire periphery of
brittle-willow range within the European USSR has been occupied by hybrid
plants only. "Pure" S. fragilis occurs
only eastward, i.e., east of the line connecting Vyborg, Volkhov, Uglich,
Vladimir, Arzamas, Tambov, Voronezh, Kremenchug, and Beltsy, with an
isolated locus on the Crimea Peninsula.
According to Meinshausen (1878: 314), around St. Petersburg,
S. fragilis "occurs only in
cultivation, though some plants are occasionally found here and there in
the woods, apparently as escapees from cultivation, as they are always
only staminate." Aun (1939: 43-47) remarks that in Estonia there is no
wild S. fragilis, either. All the
specimens that Aun had found blooming (in PÃĪrnu, VÃĪrru, Tartu, and Pechory
in the vicinity of Pskov) happened to be staminate. He was unable to
locate any pistillate plants despite his deliberate effort. Even local
forest rangers were unable to point to any pistillate brittle willows in
response to official requests through the Forestry Department. On the
other hand, hybrid specimens, though not too common, were encountered in
Estonia, and among those were some pistillate plants. My own observations
around Leningrad and the herbarium studies fully support the reports by
Meinshausen and Aun. I conclude that in Estonia and around Leningrad, the
distribution of brittle willow is similar to what is observed around
Moscow. The same refers to Pskov and Novgorod vicinities.
I have not found opinions similar to Meinshausen's or Aun's in the
literature referring to other regions. However, according to the herbarium
analyses, the area that harbors staminate non-hybrid plants only and where
pistillate trees are lacking is actually considerably wider. It extends
westward about to the line connecting Ventspils, Vilnius, Mogilev, Kiev,
Zvenigorodka, Vinnitsa, Chernovtsy, and Kluzh. Non-hybrid specimens of
both sexes can be found only west of this line. Therefore, if at all there
are any native, primary habitats of brittle willow in Europe, it makes
sense to look for them only west of this line. Apparently this boundary
reveals the central-European distributional pattern of S. fragilis, and since the flora of central
Europe is best represented within the USSR in the Carpathians and
Transcarpathia, first and foremost one should search for primary habitats
there.
I made two attempts to find such habitats during my trips to the
Carpathians in 1957 and 1968; however, both were unsuccessful. Brittle
willow is very abundant in the Carpathians. Occasionally (e.g., along a
small river named Khustinka), I encountered populations with considerable
individual variability among plants that appeared to be the "pure"
species, â an evidence of normal seed reproduction. S. fragilis even occurred continuously for a few
kilometers along some small streams, such as the Lyuta River, the left
tributary of the Uzh.
However, in intact habitats where the virgin vegetation was more or
less preserved, there was evidently no room for S. fragilis. In narrow gorges, the tall forest
usually descended right to the banks of small rivers, allowing only
minimum growth of alder and some S. purpurea L. or tall herbs here and there. At the same time,
intact habitats on the banks of a large river (Tissa R.) were taken by
pure groves of S. alba, much like in
the central European Russia. There is no doubt left for me that S. fragilis is an adventive tree in the Carpathians
and Transcarpathia, though it now appears to be well naturalized at some
places there .
III. Brittle willow in Europe
Due to lack of observations and herbarium material, my analysis of
brittle willow distribution in Western Europe has to be based largely on
literature sources.
According to many respectful sources (Hjelt 1902: 86; HultÃĐn 1950:
144; Lid 1963: 251; Hylander 1966: 288), S. fragilis is an introduced
species found only in cultivation on most of Scandinavian territory,
except for extreme southern Finland, southern Sweden, and Denmark, where
it also known to escape from cultivation. In Scandinavia hybrids with S.
alba are not infrequent (described as S. viridis Fries 1928: 283). These hybrids, along with S. alba itself, also are either cultivated or
feral.
According to the author of a monograph on the British willows,
Linton (1913: 14), S. fragilis is
native in England, southern Scotland, and Wales, being introduced to
northern Scotland. According to Praeger (1934: 26), it is probably
introduced in Ireland. A more recent British source (Clapham et al. 1962:
590) depicts the same view. However, the actual situation with S. fragilis on the British Isles appears to be
quite different. Descriptions provided for this species by all
authoritative English sources (Loudon 1838: 1516; White 1890: 348; Elwes,
Henry 1913: 1754; Linton 1913: 14; Moss 1914: 17; Clapham et al. 1962:
590) clearly demonstrate that all the authors actually present hybrids
S. fragilis x S. alba under the name "S. fragilis." Characteristically, Linton (l.c.)
provided a synonym name S. russeliana
Sm. for S. fragilis. As a matter of
fact, S. russeliana has been long
recognized as a hybrid even in the western literature (Wimmer 1866: 133,
and others). The same conclusion is to be reached when one considers
identifications on herbarium specimens provided by the English.
Along with S. fragilis, the
British authors also list S. decipiens
Hoffm. Some (White 1890: 348) consider it to be a hybrid of S. fragilis and S. triandra; others (Linton 1913: 17) think of it as a subspecies
of S. fragilis. Some refrain from
interpreting this taxon (Clapham et al., l.c.). According to Linton and
the latest Flora (Linton l.c., Clapham et al. l.c.), S. decipiens appears to be found only in
cultivation, sporadically, and infrequently in Britain. Reliable
references exist only for staminate specimens. The study of Hoffmann's
authentics of S. decipiens ("S. decipiens mihi. Erlangae" LE!) as well as
material identified by the British as S. decipiens shows that all of these are actually true S. fragilis. Therefore, the situation with
S. fragilis on the British Isles is no
different from what we find around Moscow and Leningrad.
Though S. fragilis and its
hybrids are distributed all across the British Isles, there is a distinct
increase in density in the southeastern England, as it is shown in the
Atlas of the British Flora (1962: 188). This is the most continental
part of the British Isles; in those areas where the Atlantic flora
dominates, S. fragilis becomes
scarcer.
According to Coutinho (1939: 189), S. fragilis is scattered all across Portugal, both cultivated and
"nearly wild" ("subespontaneo"). However, the local Flora (Rozeira 1944)
of Tras-os-Montes, one of the largest provinces, does not list this
willow. Of those four specimens that I received from Portugal under the
name "S. fragilis," two (M. da Silva
2724; Rainha 5706) appeared to be hybrids with S. alba, and the other two had no relationship to S. fragilis (Rainha 5930âprobably a
puberulent variation of S. alba; M. da
Silva 2432âS. babylonica or its
hybrid.)
In his monograph of Spanish willows, Vicioso (1951: 47), citing
Laguna (1883), provides that S. fragilis is found both wild and cultivated on most of the
territory, though it is hard to tell autochthonous specimens from
introduced, as it is always the case with plants cultivated from olden
times. Apparently the author is not at all convinced that this willow is
native in Spain. Merino (1906: 616) states that S. fragilis is definitely only a cultivated
plant in Galicia. The description and illustration in Vicioso's monograph
do not depict all characteristic features of S. fragilis. Most probably, hybrids S. fragilis x S. alba have
been actually described in Spain as S. fragilis. Remarkably, GÃķrz (1929: 15) believed that S. fragilis does not grow on the Iberian
Peninsula, but is substituted there by S. neotricha Goerz, which resembles S. australior Anderss. (= S. excelsa Gmel.) from the Caucasus. Obviously, S. neotricha is yet another designation for the
same hybrid cycle S. fragilis x
S. alba. In any case, there is no talk
about autochthonous S. fragilis
existing on the Iberian Peninsula.
Major sources report S. fragilis
to be distributed across "nearly the entire" France (Camus 1904: 76; Coste
1906: 271; Rouy 1910: 193); however, according to Bonnier (s. a.; ca.
1930: 40), S. fragilis is not found in
western, southwestern, and Mediterranean France. Neumann, a contemporary
Austrian authority on willows, believes, mostly basing on own
observations, that S. fragilis no
longer grows around Paris (Neumann in litt. a. 1962). According to
Chassagne (1956: 260), in the French Massif Central S. fragilis is often cultivated, and it is hard
to tell whether it is growing there naturally. Chassagne believes that the
species has an oriental origin and advances due to cultivation. Thus we
can hardly suggest that S. fragilis is
aboriginal in France. As for hybrids with S. alba, they are not infrequent. According to Chassagne (1956:
259), these hybrids are found even more often than the parental
species.
Buser (1940: 629) considered S. fragilis "by far, the rarest of all willows" in Switzerland.
According to Buser, "true S. fragilis
... is just as rare, as hybrids S. alba x S. fragilis are
common in Swiss piedmont. So it is not surprising that the Swiss have been
taking hybrids for the species or at least have not been distinguishing
between the species and hybrids... Nearly all of "S. fragilis" in local Swiss "floras" are
actually hybrids. This situation can only be explained if we admit that
S. fragilis as well as its hybrids are
not native, but rather introduced" (Buser 1940: 631). The recent Flora of Switzerland (Hess et al. 1967: 666), which includes Tirol, the French
Jura Mts., and Savoy, treats S. fragilis as an introduced species within the entire
area.
The Italian researcher of the 19th century Bertoloni (1854: 303)
reported S. fragilis for Palermo only.
Later authors believed it was scattered all across Italy (Parlatore 1867:
220; Borzi 1885: 138; Fiori 1923: 341). However, local "floras" portrait
S. fragilis as a random alien species
that shows no affinity to any particular region or native plant community.
Thus in Verona this "rare, random" tree is known from a single location
(Goiran 1898: 19). For Triest (Marchesetti 1896-97), Reggio-Emilia and
Modena (Negodi 1944), as well as the Silagian Mts. (or La Sila) in
Calabria (Sarfatti 1959) S. fragilis
is not at all listed. It is known from just a single locality in the Roman
Apennines (Zangheri 1966: 86) and a single place within Teramo Province
(Zodda 1954: 66). For the Sicily (Lojacono 1904: 392), it is only the
hybrid S. alba x S fragilis that is mentioned; besides, the vague
description makes one suspect that forms of S. alba are described instead. There are more examples like this.
Hence in Italy S. fragilis is not
aboriginal, either.
In Austria and Germany most authors interpret S. fragilis as a common species growing in
natural habitats. However, in these countries, as well as in Britain,
Switzerland, and others, hybrids of S. fragilis and S. alba have
habitually been taken for S. fragilis.
A series of hybrid forms have been described under binary names: S. rubens Schrank 1789: 226; S. fragilissima Host 1828: 6; S. palustris Host 1828: 7; S. excelsior Host 1828: 8. Even though Wimmer
(1866: 133-134) had pointed to their hybrid nature, K. Koch, an authority
in German dendrology, included them all in S. fragilis (1872: 514). The German-Swiss Synopsis (W. Koch
1907: 2300), similarly to the English sources, designated the true,
non-hybrid S. fragilis as "var.
decipiens," while noting that it was
known perhaps only in cultivation. Hartig (1851: 563) remarked that
S. russeliana Sm. (that is, a hybrid)
was common in northern Germany, while non-hybrid S. fragilis was so rare there that he had only
seen it in botanic gardens. According to Toepffer (1914: 191), in Bavaria,
all records for S. fragilis needed
revision due to constant mixing with hybrid plants. As per Toepffer, the
true S. fragilis (which he as well
named "var. decipiens") was rare in
Bavaria. In Wurttemberg, according to Bertsch (1951: 46), only hybrids
were encountered, all of them originating from cultivated specimens. In
the Northern German Lowlands, non-hybrid S. fragilis was reported rare and even completely missing from
many areas (GÃķrz 1922: 31). Neumann (in litt.) also believes that
S. fragilis is not present in the
western part of Northern German Lowlands and beyond, in the
Netherlands.
During the fall of 1964, I found some small scattered non-hybrid
S. fragilis growing along small
streams in the Gartz southern piedmont. These habitats appeared to be
completely similar to those occupied by this willow within this country:
in the Carpathians and even around Smolensk and Moscow. I also found
hybrids in-between Halle and Jena. The herbarium material from Germany that I
had at my disposal contained a number of non-hybrid as well as hybrid
specimens of both sexes. I concluded that within the southern GDR and
central BRG the situation with brittle willow is similar to that in the
western regions of our country, that is, the tree has been nearly
completely naturalized, so that occasionally it appears as a native. At
the same time, in the northwest and southwest the status of brittle willow
is closer to that in France and England.
In Lower Austria, S. fragilis
used to be considered as "one of the most common willows, particularly in
the Danube Valley" (Kerner 1860: 185). However, later on Rechinger (1957:
67) remarked that all of S. fragilis
from the Danube were actually hybrids.
In Belgium, S. fragilis has been
known as mostly cultivated, its hybrids encountered much more often than
the species (LawalrÃĐe 1952: 36).
In Poland, Czechoslovakia, Hungary, and Romania, S. fragilis appears to be frequent. It is
usually mentioned in "floras" as a native tree. However, it is hardly
possible that the origin of S. fragilis in Romania and Czechoslovakia could be drastically
different from its origin in our Carpathians; or else that the situation
in Poland fundamentally differs from what takes place in Germany or the
Baltic States. Even though non-hybrid plants of both sexes are not
infrequent, the majority of specimens are of hybrid nature, as per
WoÅoszczak (1889: 292), for Transylvania or Szafer (1921: 33), for
Poland.
In Yugoslavia, S. fragilis has
been believed to be widespread (Beck von Managetta 1909: 115;
Adamoviħ 1909: 194; Rohlena 1942: 19; Å panoviÄ? 1954;
JovanoviÄ? 1955; Em 1967; and others). However, a note by Hirc (1904:
158) remarkably testifies for absence of herbarium collections from
Croatia. S. fragilis is also characteristically absent from remote ravines
in Macedonia, where the flora should still be most primeval (SoÅĄka
1938-39). Those few specimens from southern Yugoslavia that I examined
were all hybrids.
Velchev (1966: 56) reports S. fragilis being distributed in Bulgaria "across the entire
country." However, the description that he provides rather fits hybrids.
According to Stefanov (1943), S. fragilis in Bulgaria is a boreal element restricted to the
northwest. According to Chernyavskiy et al. (1959: 94), stands of
S. fragilis in Bulgaria are of
"secondary origin." Regretfully, this statement is elaborated no
further.
In Greece S. fragilis apparently
is completely absent.
While summarizing data on S. fragilis range in Europe outside the Soviet Union, we can
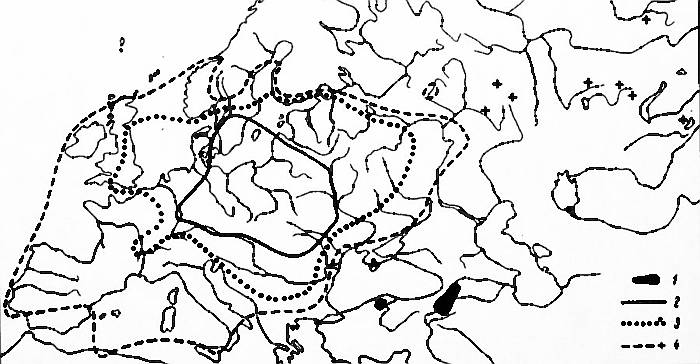
distinguish the same three zones as within the Soviet Union (Fig.
1):
* core zone where S. fragilis
has been nearly entirely naturalized and is represented by non-hybrid
trees of both sexes;
* transitional zone, where the species is still quite common,
though among non-hybrid plants staminate ones overwhelmingly
dominate;
* peripheral zone, where virtually only hybrids are represented,
being scattered, sporadic, or cultivated only.
Of course the demarcation lines are at this point very much
approximate. For the refinement, much more information is needed than I
have had at my disposal.
Fig. 1. Distribution of Salix fragilis L.
1. primary range
2.
area where S. fragilis is nearly
naturalized, non-hybrid plants of both sexes encountered
3. distribution limits for non-hybrid staminate plants, their
reproduction spontaneous, vegetative
4. boundary of area where nearly
exclusively hybrid plants are encountered, originating from
plantings
Literature sources also report a rather large-scale introduction of
S. fragilis to North America, where it
appears to be also nearly naturalized. All those specimens from North
America that I have examined happened to be only hybrids.
IV. Brittle willow in the Near East, Asia Minor, and the
Caucasus
In eastern Asia Minor, southern and eastern Transcaucasia, Iran, and
the Soviet Central Asia there is a common, widely distributed willow,
S. excelsa Gmel. This species is a
race of S. alba, closely related to
it, though distinct in a number of characters that resemble S. fragilis: non-weeping crown, stout, brittle
branches, stout buds, less pubescent leaves and branchlets, and more
pubescent bracts. It was first described from northern Iran in 1774 by
Gmelin, the younger, but then forgotten and described for the second time
by Andersson (1867: 43) as S. australior Anderss. or S. fragilis Îī australis
(Andersson 1868: 210). Andersson placed this tree in the S. fragilis cycle by mistake and then suggested
that the European S. fragilis had
originated from southwest Asia (Andersson 1867: 41).
Relying on Andersson's works, Boissier (1879: 1184) also listed
S. fragilis for Syria, Asia Minor, and
Transcaucasia. However, it soon became evident that S. australior Anderss. was not at all a form of
S. fragilis, but rather belonged to
S. alba cycle. Despite this
clarification, due to the authority of Boissier, almost until recently
S. fragilis has been considered an
"oriental" species present in Syria and Lebanon (Bouloumoy 1930: 316; Post
1933: 530); Jordan (NÃĄbÃĐlek 1929: 24); Iran (Parsa 1950: 1351); western
and southern Transcaucasia (Medvedev 1883: 228, Nazarov 1936: 203);
Dagestan (Lvov 1956: 77); the western and eastern Caucasus (Makhatadze
1961: 51); and the Russian Central Asia (Fedchenko 1915: 295, Protopopov
1953: 36). There is no doubt that all these listings were based on either
erroneous identifications or uncritical borrowing.
Meanwhile, the true S. fragilis
was actually discovered growing in the East. It was quite wild, neither
adventive, nor deliberately introduced, its populations represented by
both sexes. Besides, hybridization with S. alba was completely absent or at least very insignificant. It
was first collected in 1892 by Sintenis; however, the first reliable
record of S. fragilis in Asia Minor
belongs to GÃķrz (1930: 113). He discovered specimens collected by S.
Turkevich in former Kars [Province of Turkey; Russian Turkestan at the
time of collection]. Even though GÃķrz considered this find "amazing, " he
still under-evaluated its importance. It did not even occur to him to
re-examine the collections he had at hand. Indeed he failed to correctly
identify some true S. fragilis that he
collected himself.
I examined the following specimens of S. fragilis from its eastern range:
1. Paphlagonia, wilajet Kastambuli, Tossia, Utsch-tschesme,
3.VI.1892, P. Sintenis No. 4089, fl. femin. et fol.;
2. districtus GÞmÞsch-hane, supra Bayburt, 1500 m, frutex inter
stirpes typicas S. australioris idque
prorsus sponte nascens, 8.VI.1931. R. GÃķrz (Sal. Asiat. No. 37: "S. australior var. pseudofragilis Goerz var. nova"), fol.;
3. inter GÞmÞsch-hane er Bayburt, 1600 m, sponte nascens,
16.VI.1931. R. GÃķrz (Sal. Asiat. No. 37-a: "S. australior ad var. pseudofragilem Goerz vergens"), fl.
femin.;
4. Prov. Kastamonu, 10 km from Daday to Effani, 900-1000 m,
stream-side in pinetum, 30.VII.1962. Coode et Yaltirik No. 38610,
fol.;
5. Prov. Kastamonu, between Seydiler and Kure, 1200 m, salix scrub
near stream, 30.VII.1962, P. Davis No. 38464, fol.;
6. Karsskaya obl., bliz st. Promezhutochnoi, bereg r. Kyaklik [Kars
Oblast, close to Promezhutochnaya Station, bank of the Kyaklik
River], 11.V.1914, S. Turkevich No. 245, fl. femin., masc.;
7. Karsskaya obl., mezhdu Sarykamyshem i Karakurtom v Tyaklitskoi
balke [Kars Oblast, between Sarikamis and Karakurt in the Tiyaklit
Depression], T. Roop, fol.;
8. Sanjak Erzerum, tugai r. Khnys-chai bliz Khnyskaly [Erzurum
Province, Hinisçayi River forested floodplain near Hiniskaly],
14.VII.1916, V. Sapozhnikov and B. Shishkin, fol.;
9. bliz Akhaltsikha, r. Poshov-chai [near Akhaltsikhe, the
Posof-çayi River], 9.VIII.1920, D. Sosnovskiy, fol.
The latter is the only sample known so far from the USSR
territory.
There is no indication that any of the listed specimens originate
from cultivated plants. Accompanying hybrid series, which are so common in
European collections, are absent here. Indeed hybrids (T. Roop 1911, LE)
were collected only around the Kars Fortress, from the landscape that had
been greatly disturbed for a long time.
According to Grossheim (1945: 27), S. fragilis is commonly cultivated in the Northern Caucasus. If
this is true (I did not see any samples), then S. fragilis (or, more likely, its hybrids) apparently had been
brought to the Northern Caucasus and Siberia with migrations of the
Russian and Ukranian population from the west and northwest rather than
directly from Asia Minor.
Conclusions
Thus from northern and northwestern Asia Minor, where it had a small
primary range in the mountains, brittle willow was somehow introduced to
Europe, where it advanced primarily by means of vegetative reproduction
and formed an enormous secondary range. The plant has been nearly
completely naturalized in the central part of this range; its
alien nature is more obvious in the peripheral
parts. Across all of its secondary range, S. fragilis hybridizes with S. alba on a large scale; in significant parts of the range,
particularly peripheral, non-hybrid specimens are completely
absent.
When and where exactly in Europe did S. fragilis first appear? Is it still continuing to advance? One
might assume that it appeared first within the center of its present
range. However, the actual events could as well be quite different: it is
also possible that the formation of the present locus has been influenced
by climatic factors rather than historic. S. fragilis could have entered Europe somewhere at a peripheral
part of its modern range, such as Italy or the Balkans. In any case,
S. fragilis was likely to be promptly
transported over a great distance. As for its arrival on the Russian
territory, it is clear enough that it came from the west rather than
across the Caucasus.
Quite a number of S. fragilis
herbarium samples have been preserved for 100-150 years. They provide
evidence for the conclusion that during the recent 100-150 years there has
been no significant dynamics in the distribution of this species. For
example, non-hybrid staminate and hybrid pistillate specimens were
collected around Moscow, respectively, in 1837 and 1823; around St.
Petersburg, respectively, in 1850 and 1827; non-hybrid specimens of both
sexes were already present around Mogilev in 1862, and so on. Apparently
the process of colonization in Europe has essentially been completed, so
that S. fragilis has attained its
climatically determined limits. Certainly this is as well indicated by the
concentric zoning pattern described above. Had the process of colonization
been continuing actively into the present time, such a structure could not
yet have been established. Non-hybrid plants definitely have a greater
capacity for self-dispersal than hybrids. Hybrids continue to expand their
range only through new plantings. From the economic standpoint, hybrids
are more valuable, as they grow better. Additionally, hybrids might be
more tolerant to climatic conditions of Mediterranean Region as well as
those in Siberia, since one of the parental species, S. alba, grows naturally in both these regions.
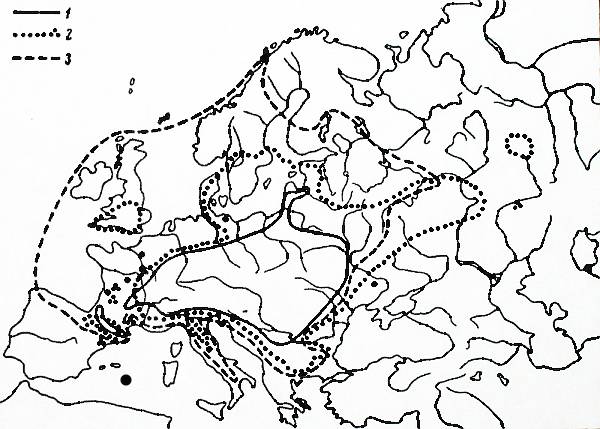
The similarity of the secondary range of S. fragilis and natural ranges of a few European species (Fig. 2)
also signifies that climatic limits might have been already
reached.
Fig. 2. Areas of some European species that resemble
S. fragilis area (adapted
from Meusel et al. 1965)
1. Carex brizoides Juslen. 2. Carex montana L. 3. Anemone nemorosa L.
From the time of the Turkish conquest of the eastern Byzantine
provinces in the 11-12th centuries and to the mid-19th century there was
literally no connection between Europe and eastern Asia Minor. Hence it is
safe to speculate that S. fragilis was
introduced to Europe during the Byzantine Empire rule or maybe even during
the Roman epoch. In one of local British "floras" (Grose 1957: 508), I
came across some curious evidence. In ancient Saxon land deeds of
Wiltshire County (there are fifteen of them, the oldest dated 796), some
willows ("withies") are mentioned as markers of property boundaries.
S. fragilis presently grows at nearly
all of the named locations. Boundary markers were of course preserved and
restored; therefore, it is not improbable that the willows growing there
today are direct descendants of trees that were planted more than a
thousand years ago.
Classic examples of ginkgo, metasequoia, and franklinia show that a
plant with an extremely narrow relict range can be very viable, highly adaptable,
and tolerant to a wide range of conditions. Hence a narrow primary range
of S. fragilis should not be
considered an obstacle for this willow to attain a wide secondary range.
Besides, the phytoclimate of the forested regions in eastern Asia Minor
has much in common with the climate of central Europe.
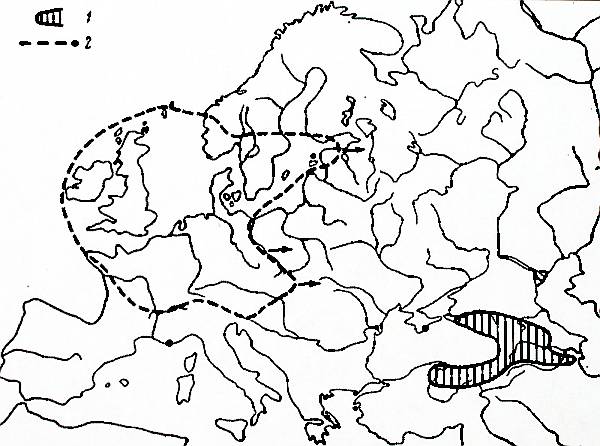
Veronica filiformis Sm. provides
another example of a species with the natural range in the Caucasus and
Asia Minor extensively naturalized in Europe. Much has been said about it
(Lenmann 1942; Thaler 1953; KornaÅ, Kuc 1954; Bangerter, Kent 1965;
Jehlik, Slavik 1967). This veronica was first found in the wild in 1893 in
France. It has continued colonizing the European countries since then,
mostly (or perhaps exclusively) vegetatively. Its present range embraces
nearly all of central Europe (Fig. 3). Recently it has been found in
Estonia (Eichwald 1960). Similarly to S. fragilis, it appears to advance to the Russian territory from
the west rather than south, i.e., not directly from the Caucasus.
Fig. 3. Distribution of Veronica filiformis Sm. (generalized)
1. primary range 2. secondary range
There remains a single question concerning S. fragilis that yet awaits an answer. Why do
staminate non-hybrid plants have a wider range than pistillate
non-hybrids? Perhaps those were only staminate plants that were initially
introduced. Pistillate plants then might have appeared only many
generations later, as a result of hybrid segregation. Or else, staminate
specimens might have been more capable of vegetative regeneration. It is
quite possible that the right answer is none of the above. It has yet to
be discovered.
References
Adamoviħ L.
1909. Flora Serbiae austro-orientalis. Salix. Rada
Jugosl. Akad., 177: 194-195.
Andersson N. J.
1867. Monographia Salicum 1. Kongl. Svenska
Vetens.-Akad. Handl. 6(1). Stockholm.
Andersson N. J.
1868. Salix. In A.P. De Candolle (ed.).
Prodromus systematis naturalis regni vegetabilis 16
(2): 190-323. GenÃĻve.
Atlas of the British Flora. F.H. Perring, S.M. Walters (eds.).
1962. LondonâEdinburgh.
Aun K.
1939. Eesti pajud. Tallinn.
Bakunin A.A.
1879. Spisok tsvetkovykh rasteniy Tverskoi flory [Flowering
Plants of the Tver Flora. A Checklist]. Tr. SPb obshch.
yestestvoisp. 10.
Bangerter E.B., Kent H.D.
1965. Additional notes on Veronica filiformis. Proc. Bot. Soc. Brit. Isles
6.
Beck v. Mannagetta G.
1909. Flora von Bosnien ... Salicaceae. Wiss. Mitt.
aus Bosnien und Hercegovina 9: 114-119. Wien.
Bertoloni A.
1854. Flora italica 10.
Bononiae.
Bertsch K.
1951. Jahresh. Vereins VaterlÃĪnd. Naturk.
WÞrttemberg 106: 46 (fide LawalrÃĐe 1952: 38; non
vidi).
Boissier E.
1879. Flora Orientalis 4.
GenÃĻveâBasel.
Bonnier G. (s.a.; ca.
1930). Flora complÃĻte illustrÃĐe 10.
Paris.
Borzi A.
1885. Compendio della flora forestale
italiana. Messina.
Bouloumoy L.
1930. Flore du Liban et de la Syrie.
Paris.
Buser R.
1940. Kritische BeitrÃĪge zur Kenntnis der schweizarischen
Weiden. Ber. Schweiz. Bot. Ges. 50:
567-788.
Camus A., Camus E.-G.
1904. Classification des saules d'Europe et
Monographie des saules de France 1. Paris.
Chassagne M.
1956. Inventaire analytique de la flore d'Auvergne et
contrÃĐes limitrophes 1. Paris.
Chernyavskiy P.I., Ploshchakova L. Nedyalkova S., Dimitrov I.
1959. Drveta i khrasti v gorite na Blgariya [BÃĪumer
und StrÃĪucher in der WÃĪldern Bulgariens].
Sofiya
Clapham A.K., Tutin T.G., Warburg E.F.
1962. Flora of the British Isles. 2nd ed.
Cambridge.
Coste H.
1906. Flore descriptive et illustrÃĐe de la
France 3. Paris.
Coutinho A.X.P.
1939. Flora de Portugal. 2nd ed.
LisbÃĩa.
Eichwald K.
1960. Niitjas mailaneâEesti rohumaid ohustav umbrohutaim.
Eesti Loodus 3.
Elwes H.J., Henry A.
1913. Trees and shrubs of Great Britain and
Ireland. 7. Edinburgh.
Em H.
1967. Pregled na dendroflorata na
Macedonija. Skopje.
Fedchenko B.A.
1915. Rastitelnost Turkestana [Vegetation
of Turkestan.] Petrograd.
Fiori A.
1923. Nuova flora analitica d'Italia 1.
Firenze.
Flerov A.F.
1902. Flora Vladimirskoy gub. [Flora of
Vladimir Government]. Moscow.
Flerov A.F.
1912. Flora Kaluzhskoy gub. [Flora of
Kaluga Government] Kaluga.
Fries E.M.
1828. Novitiae florae Suecicae. 2nd ed.
Lund.
GÃķrz R.
1922. Ãber norddeutsche Weiden. Feddes
Repert. 13.
GÃķrz R.
1929. Les saules de Catalogne.
Cavanillesiae 2, 7-10. Barcelona.
GÃķrz R,
1930. BeitrÃĪge zur flora Kleinasiens. Salix. Feddes
Repert. 28.
Goiran A.
1898. Juglandaceae et
Salicaceae veronenses. Bull. Soc. bot.
Ital. 18-24.
Grose J.D.
1957. Flora of Wiltshire.
Devizes.
Grossheim A.A.
1945. Flora Kavkaza [Flora of the Caucasus]
3. 2nd ed. Baku.
Hartig T.
1851. VollstÃĪndige Naturgeschichte der forstlichen
Kulturpflanzen Deutschlands. Berlin.
Hess H.E., Landolt E., Hirzel R.
1967. Flora des Schweiz. 1.
BaselâStuttgart.
Hirc D.
1904. Revizija Hrvatske flore. Salix. Rada Jugoslav.
Acad. 159: 157-162.
Hjelt H.
1902. Conspectus florae fennicae. 2(1).
Helsingforsiae.
Hoffmann G.F.
1791. Historia Salicum iconibus illustrata
2, fasc. 1. Lipsiae.
Host N.T.
1828. Salix.
Vindobonae.
HultÃĐn E.
1950. Atlas Ãķver vÃĪxternas utbredning i
Norden. Stockholm.
Hylander N.
1966. Nordisk KÃĪrlvÃĪxtflora 2.
Stockholm.
Jehlik, N., Slavik B.
1967. DoplÅky k rozÅĄireni Veronica filiformis v Äeskoslovensku.
Preslia 39.
JovanoviÄ? B
1955. Sumske fitocenoze i stamista Suve Planine.
Glasnik sumarskog fakult. 9.
Beograd.
Kerner A.
1860. NiederÃķsterreichische Weiden. Verhandl.
zool.-bot. Ges. Wien 10.
Koch K.
1872. Dendrologie 2 (1).
Erlangen.
Koch W.D.J.
1907. Synopsis der Deutschen und Schweizer
Flora 3. Aufl. Hersg. E. Hallier u. A. Brand.
Leipzig.
KornaÅ J., Kuc M.
1954. Veronica filiformis
Sm. â nowy we flore polskiej uciaÅžliwy chwast lakowy.
Fragm. flor. et geobot. 1(1).
Laguna M.
1883. Flora forestal EspaÃąola 1. Madrid
(n.v.)
LawalrÃĐe A.
1952. Salix. In
Flora gÃĐnÃĐrale de Belgique 1(1).
Bruxelles.
Lehmann E.
1942. Die EinbÞrgerung von Veronica filiformis in Westeuropa.
Gartenbauwissenschaft 16.
Lid J.
1963. Norsk og Swensk flora.
Oslo.
Linton E.F.
1913. The British willows. Suppl. to
J. Bot. 51.
Lojacono M.
1904. Flora Sicula 2(2).
Palermo.
Loudon J.C.
1838. Arboretum et frutecetum Britannicum
3. London.
Lvov P.L.
1956. Opredelitel glavneyshikh derevyev i kustarnikov
Dagestana [Guide to Most Important Trees and Shrubs of
Dagestan.] Makhachkala.
Makhatadze L.B.
1961. Salix. In
Dendroflora Kavkaza [Dendroflora of the Caucasus]
2. Tbilisi.
Marchesetti C.
1897. Flora di Trieste.
Trieste.
Mayevskiy P.F.
1892. Flora Sredney Rossii [Flora of
Central Russia], 1st ed. Moscow;
Mayevskiy P.F.
1895. Flora Sredney Rossii [Flora of
Central Russia], 2nd ed., S.I. Korzhinskiy ed. Moscow.
Mayevskiy P.F.
1902. Flora Sredney Rossii [Flora of
Central Russia], 3rd ed., B.A. Fedchenko ed. Moscow.
Mayevskiy P.F.
1912. Flora Sredney Rossii [Flora of
Central Russia], 4th ed., D.I. Litvinov ed. Moscow.
Medvedev Ya.S.
1883. Derevya i kustarniki Kavkaza [Trees
and Shrubs of the Caucasus]. Tiflis.
Meinshausen K.F.
1878. Flora Ingrica. St.
Petersburg.
Merino B.
1906. Flora descriptiva e ilustrada de
Galicia 2. Santiago.
Meusel H., JÃĪger E., Weinert E.
1965. Vergleichende Chorologie der zentraleuropÃĪischen
Flora 1. Jena
Moss C. E.
1914. The Cambridge British Flora 2.
Cambridge.
NÃĄbÃĐlek F.
1929. Iter turcico-persicum 4. Spisy prirodoved. fak.
univ. Brno 105.
Nazarov [Nasarov, Nasaroff] M.I.
1936. Salix. In
Flora SSSR [Flora of the USSR] 5.
MoscowâLeningrad.
Nazarov [Nasarov, Nasaroff] M.I.
1949. Salix. In
Flora Belorussii [Flora of Byelorussia] 2.
Minsk.
Negodi G.
1944. Flora delle provincia di Modena e Reggio Emilia.
Atti Soc. Natur. Moden. 75.
Nevskiy M.L.
1952. Flora Kalininskoy oblasti [Flora of
Kalinin Oblast] 2. Kalinin.
Parlatore F.
1867. Flora Italiana 4.
Firenze.
Parsa A.
1950. Flore de l'Iran 4.
Teheran.
Penkovskiy V.M.
1901. Derevya i kustarniki, kak dikorastushchiye, tak
i razvodimyye [Wild and Cultivated Trees and Shrubs.]
Kherson.
Petunnikov A.N.
1901. Salix. In [Critical Review of Moscow Flora. III.]
Tr. SPb obshch. yestestvoisp. 31(3):
18-40.
Post G.E.
1933. Flora of Syria, Palestina, and Sinai
2. 2nd ed. Beirut.
Praeger R.L.
1934. The botanist in Ireland.
Dublin.
Pravdin L.F.
1951. [Willow.] In Derevya i kustarniki
SSSR [Trees and Shrubs of the USSR] 2.
MoscowâLeningrad.
Protopopov G.F.
1953. Salix. In
Flora Kirgizskoy SSR [Flora of Kirghiz SSR] 4:
9-37. Frunze.
Rechinger K.H.
1957. Salix. In Hegi G.
Illustrierte Flora von Mitteleuropa 3(1): 44-135.
Aufl. 2. MÞnchen.
Rohlena J.
1942. Conspectus florae Montenegrinae.
Preslia 20-21.
Rouy G.
1910. Salix. In
Flore de France 12: 189-252. Paris.
Rozeira A.
1944. A flora da provincia de Tras-os-Montes. Memor.
Soc. Broter. 3.
Rozen V.V.
1916. [Checklist of Plants Found in Tula Government up until
1916.] Izv. Tulsk. obshch. lyubit. yestestvozn.
4.
Sarfatti G.
1959. Prodromo della flora della Sila.
Webbia 15.
Schrank F.P.
1789. Baierische Flora 1.
MÞnchen.
Seemen O., v.
1908-1910. Salix. In Ascherson
P., Graebner P. Synopsis der mitteleuropÃĪischen
Flora 4: 54-350. Leipzig.
Siuzev P.V.
1901. [On some new plants of Moscow flora.] Tr. bot.
sada Yuryev. un-ta 2(1).
Skvortsov A.K.
1956. [Some additions and amendments to the willow flora of West
Siberia.] Sistem. zametki gerbariya Tomsk. un-ta
79-80: 13-15.
Skvortsov A.K.
1966. [Review of willows from the Caucasus and Asia Minor.]
Tr. Bot. in-ta AN ArmSSR 15: 91-141.
Skvortsov A.K.
1968. Ivy SSSR [Willows of the USSR.]
MoscowâLeningrad.
SoÅĄka T.
1938-1939. BeitrÃĪge zur Kenntnis der Schluchtenfloren von SÞdserbien
1-7. Glasnik Skopskog nauchnog drushtva 18,
20.
Å panoviÄ? T.
1954. Vrbe naÅĄih podunavskih ritova.
Å umarski List 78: 506-521.
Zagreb.
Stefanov B.
1943. Fitogeografski elementi v Blgaria. Sb. Blg.
Akad. 39. Sofia.
Syreishchikov D.P.
1907. Illustrirovannaya flora Moskovskoy
gub. [Illustrated Flora of Moscow Government] 2.
Moscow.
Syreishchikov D.P.
1927. Opredelitel rasteniy Moskovskoy gub.
[Guide to the Plants of Moscow Government.] Moscow.
Szafer W.
1921. Salicaceae. In Flora
Polska 2: 24-47. KrakÃģw
Thaler I.
1953. Die Ausbreitung von Veronica filiformis. Phyton 5.
Toeppfer A.
1914. Salicaceae. In Vollmann F.
Flora von Bayern: 187-202.
Stuttgart.
Tsinger V. Ya.
1886. Sbornik svedeniy o flore Srednei
Rossii [Anthology of Knowledge about the Flora of Central
Russia.] Moscow.
Vicioso C.
1951. Salicaceas de EspaÃąa.
Madrid.
White F.B.
1890. A revision of the British willows. J. Linn.
Soc. 27.
Wimmer F.
1866. Salices Europaeae.
Vratislaviae.
WoÅoszczak E.
1889. Kritische Bemerkungen Þber siebenbÞrgische Weiden.
Ãsterreich. bot. Z. 62: 291-295;
330-332.
Zangheri P.
1966. Flora e vegetazione del medio ed alto Appenino Romagnolo.
Webbia 21.
Zodda G.
1954. La flora teramana. Webbia
10.
NOTES
Translation I.Kadis
1 Jan 2008